近期,园艺学院菊花遗传育种与种质创新团队蒋甲福教授在Plant Cell & Environment发表了题为“Two B-box proteins orchestrate vegetative and reproductive growth in summer chrysanthemum”的论文,揭示了B-box蛋白CmBBX5和CmBBX8协同参与调控夏菊营养生长和生殖生长的机制。
菊花多为短日照品种,于秋季开放。夏菊属于光周期不敏感类型,在长短日照下均可开花,但目前品种较少,开花机理也不清楚。团队前期发现CmFTL1是夏菊“优香”在长日照条件下的开花素,且被B-box蛋白CmBBX8直接转录调控,说明该BBX8-FT模块在夏菊开花调控中发挥重要作用(Plant Biotechnology Journal 2020);CmBBX8同一亚家族成员CmBBX7在调控夏菊开花中也发挥了重要作用,和CmBBX8协同促进CmFTL1的表达,在加速开花过程中起着“涡轮增压”的作用(Molecular Horticulture 2023)。

但夏菊如何防止过早开花,保证正常营养生长的呢?首先利用CmBBX8酵母双杂交筛库,然后通过体内体外实验确认CmBBX5与CmBBX8的互作。组织表达模式分析表明,CmBBX5在营养生长时期各组织的表达水平高于CmBBX8,而在生殖生长时期各组织的表达水平低于CmBBX8。为了明确CmBBX5基因的功能,创制超表达和融合抑制基因沉默菊花株系。相比较野生型,超表达株系推迟7-10 天开花,融合抑制沉默株系早花6-11天。通过酵母单杂交实验发现CmBBX5不结合CmFTL1的启动子,但EMSA实验发现CmBBX5会减弱CmBBX8结合CmFTL1的能力,同时荧光素酶实验表明CmBBX5会抑制CmBBX8促进CmFTL1表达的能力;通过原生质体瞬时转化实验发现,CmBBX5不影响CmBBX8的转录活性(图1)。为了进一步明确CmBBX5与CmBBX8的互作机理,通过双基因遗传转化菊花,证明CmBBX5部分依赖CmBBX8调控夏菊开花。
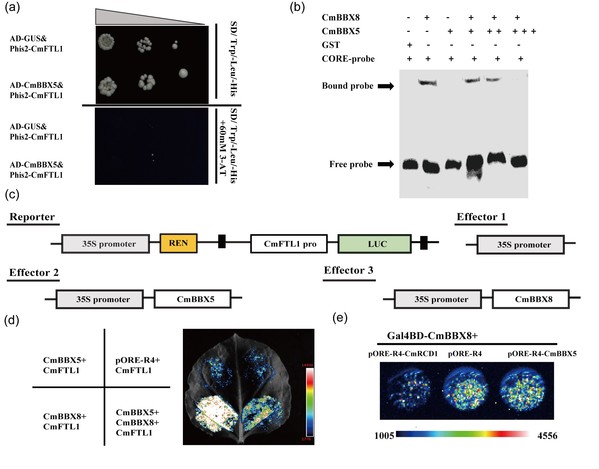
图1 CmBBX5抑制CmBBX8与CmFTL1启动子之间的相互作用以及对CmBBX8转录活性影响
团队前期发现,夏菊中CmRCD1通过抑制CmBBX8的转录活性及其结合CmFTL1的启动子能力抑制开花(Horticulture Research 2021),CmERF3通过抑制CmBBX8的转录活性来抑制开花(Plant Cell and Environment 2023),该研究中CmBBX5通过影响CmBBX8与CmFTL1启动子结合能力来抑制开花(图2)。由此可见,夏菊“优香”设置了多种开花“刹车”方式,以防止在不成熟时过早开花导致的品质不良。因此,每个生命都有生长规律,需要在合适的时间做合适的事情。
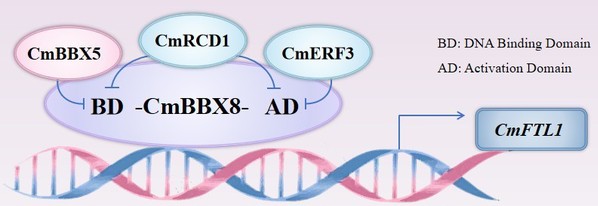
图2 CmBBX5/CmRCD1/CmERF3与CmBBX8调控开花模型
园艺学院博士生王琦为论文第一作者,蒋甲福教授为该论文的通讯作者,周李杰讲师、陈素梅教授和陈发棣教授参与了此项工作的研究。该研究得到了国家自然科学基金、中央高校基本研究基金和江苏省高等学校优势学科项目资助。
园艺学院陈发棣教授领衔的菊花遗传育种与种质创新团队针对菊花开花时间调控机制不明,围绕光温和激素调控开花优异基因挖掘与分子改良开展了较为深入的研究,目前已在Plant Biotechnology Journal、Plant Physiology、The Plant Journal、Plant Cell and Environment、Horticulture Research、Journal of Experimental Botany和Molecular Horticulture等期刊发表了多篇研究论文。
文章链接:http://doi.org/10.1111/pce.14919

